Barsboldia
Vois les stats, capacités et rôle de Barsboldia sicinskii sur notre serveur survie.
Barsboldia vivait au Crétacé supérieur, durant le Maastrichtien inférieur, il y a environ 72 à 70 millions d'années. Cette période correspond à l'un des derniers chapitres de l'ère des dinosaures non-aviens, quelques millions d'années avant l'extinction massive du Crétacé-Paléogène. La Formation Nemegt, dans la région d'Ömnögovi au sud de la Mongolie, était alors une vaste plaine alluviale traversée de rivières, abritant l'un des écosystèmes les plus diversifiés du Crétacé asiatique. Barsboldia partageait ce milieu avec des contemporains imposants : le tyrannosaure géant Tarbosaurus bataar, le hadrosauridé Saurolophus angustirostris, l'énigmatique Deinocheirus mirificus et le thérizenosauridé Therizinosaurus cheloniformis.
Barsboldia sicinskii a été décrit scientifiquement en 1981 par les paléontologues polonaises Teresa Maryańska et Halszka Osmólska, deux figures majeures des expéditions polono-mongoles des années 1960-1970. Le genre est dédié à Rinchen Barsbold, paléontologue mongol dont les travaux sur les dinosaures du Crétacé d'Asie centrale sont fondamentaux. L'épithète spécifique sicinskii honore pour sa part le géologue polonais Andrzej Siciński. Les fossiles ont été récoltés lors des expéditions conjointes polono-mongoles dans le désert de Gobi, des campagnes de terrain qui ont permis de mettre au jour une quantité remarquable de fauna du Crétacé mongol. La description originale de 1981 repose sur un matériel squelettique très partiel, ce qui a durablement compliqué la position taxonomique de cet animal.
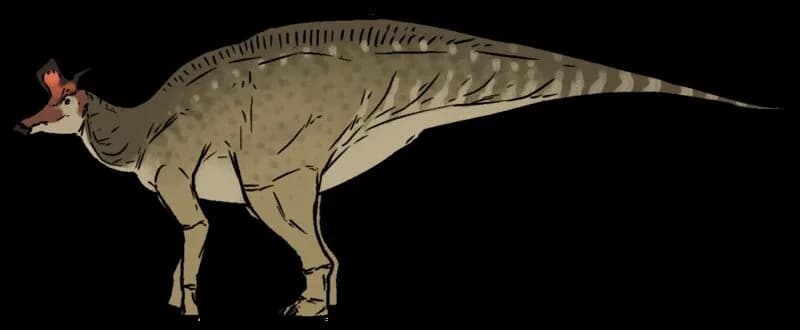
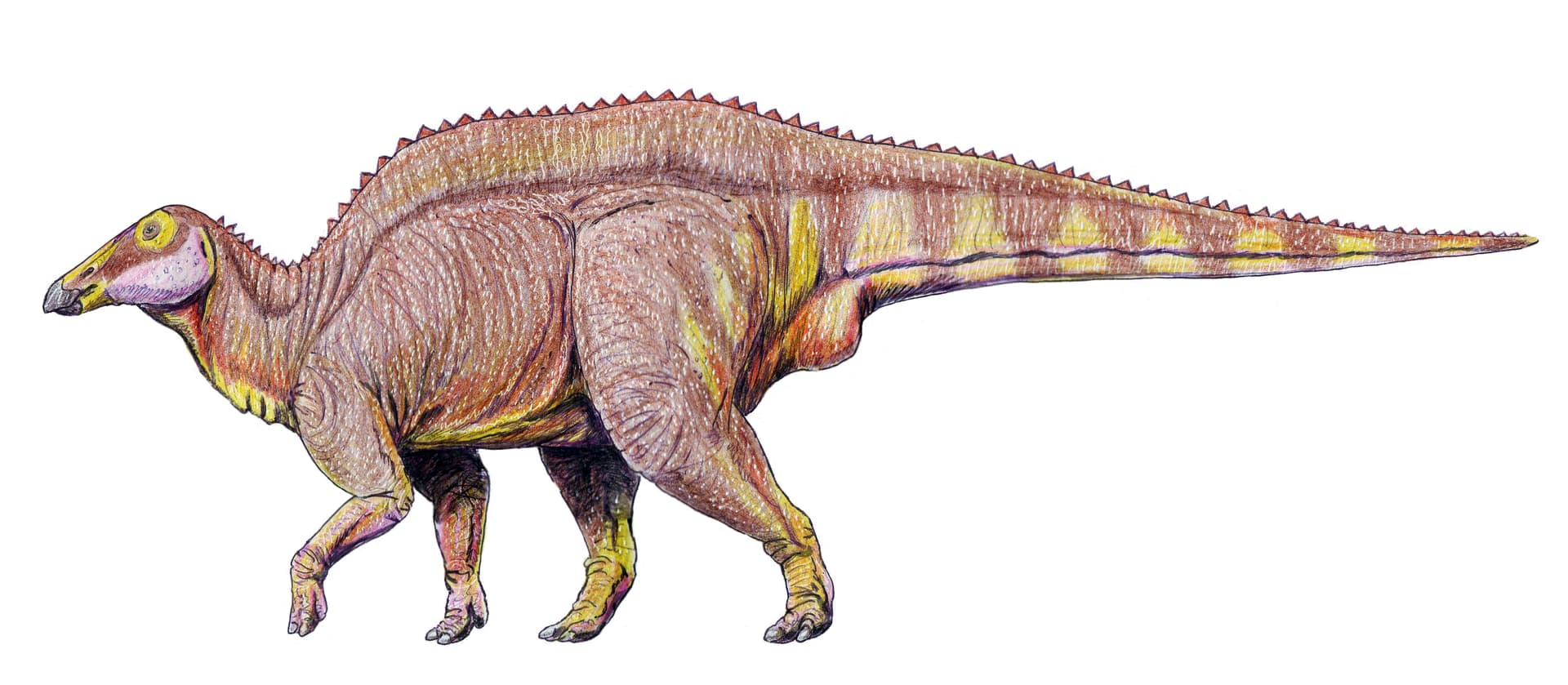
Barsboldia comptait parmi les grands hadrosauridés de la Formation Nemegt, avec une longueur estimée entre 10 et 11 mètres, certaines projections allant jusqu'à 12 mètres. Pour un hadrosauridé, ce gabarit le situait dans la même catégorie que Saurolophus angustirostris ou les grandes espèces d'Edmontosaurus. Son poids reste difficile à chiffrer précisément en raison du matériel fragmentaire disponible, mais les estimations tournent autour de 4 à 7 tonnes selon les modèles de masse corporelle appliqués aux hadrosauridés de dimensions similaires. Les épines neurales allongées des vertèbres dorsales suggèrent une architecture dorsale haute, ce qui lui donnait probablement une silhouette imposante de profil.
Comme tous les hadrosauridés, Barsboldia était un herbivore strict. Son régime alimentaire reposait sur la végétation disponible dans les plaines de la Formation Nemegt : plantes à fleurs (angiospermes), conifères, fougères et plantes basses de bord de rivière. Les hadrosauridés possédaient l'une des dentures les plus performantes du règne animal mésozoïque — des batteries dentaires composées de plusieurs centaines de dents remplacées en continu, capables de broyer des matières végétales coriaces que peu d'autres dinosaures pouvaient digérer efficacement. Chez une espèce adulte, ces batteries pouvaient compter jusqu'à 1 400 dents fonctionnelles et de remplacement empilées en colonnes verticales. Barsboldia, si son anatomie buccale correspondait au profil standard des saurolophines, se nourrissait probablement à hauteur variable, depuis le sol jusqu'à 3 ou 4 mètres de haut en position bipède. Le traitement digestif des fibres végétales était assuré par un système digestif volumineux, avec fermentation dans un intestin étendu — stratégie efficace pour extraire les nutriments de végétaux peu caloriques. La diversité végétale du Nemegt à cette époque offrait un répertoire alimentaire beaucoup plus large qu'aux hadrosauridés des formations plus arides du Crétacé asiatique.
La Formation Nemegt représente un environnement fluvial de grande échelle : des plaines inondables larges, des chenaux anastomosés, des lacs peu profonds et des zones boisées en bordure de cours d'eau. Le climat était humide et saisonnier, nettement plus tempéré que les environnements semi-arides qui caractérisent d'autres formations mongoles du Crétacé comme la Formation Djadokhta. Ces conditions favorisaient une végétation dense et une faune abondante, ce qui explique la richesse exceptionnelle du registre fossile nemegt. La coexistence avec Tarbosaurus — l'équivalent asiatique de Tyrannosaurus rex — implique que Barsboldia et les autres grands herbivores du Nemegt étaient soumis à une pression de prédation significative. Le paysage général évoque les deltas fluviaux modernes d'Asie du Sud-Est, avec une mosaïque d'habitats ouverts et forestiers.
Le matériel attribué à Barsboldia sicinskii est limité : une colonne vertébrale partielle (vertèbres dorsales et sacrées), un bassin incomplet et des fragments de côtes. Ce corpus squelettique insuffisant est la source principale des incertitudes taxonomiques persistantes autour de ce genre. La caractéristique anatomique la plus notable concerne les épines neurales des vertèbres dorsales, qui sont exceptionnellement allongées — plusieurs fois la hauteur du corps vertébral. Ces structures osseuses élevées évoquent superficiellement une voile dorsale, comme on en observe chez certains synapsides pelycosauriens ou chez le spinosaure, mais leur interprétation fonctionnelle chez Barsboldia reste incertaine : voile thermorégulée, bosse adipeuse, ou simple architecture musculaire de soutien du dos. Cette morphologie vertébrale était initialement considérée comme un caractère lambeosauriné, mais des analyses phylogénétiques ultérieures ont remis en question cette attribution.
Le comportement de Barsboldia ne peut être inféré qu'indirectement, par comparaison avec les autres hadrosauridés mieux documentés de la même époque et des mêmes milieux. Les hadrosauridés en général sont associés à des comportements grégaires — regroupements en troupeaux, déplacements collectifs, et peut-être des soins parentaux, comme le suggère le dossier fossile de Maiasaura en Amérique du Nord. Barsboldia, en tant que grand herbivore du Nemegt, coexistait avec d'autres saurolophines dont Saurolophus angustirostris, suggérant un possible partage de niche écologique ou une ségrégation alimentaire par taille ou par habitat préférentiel. La présence de Tarbosaurus comme prédateur dominant de l'écosystème indique que les grands hadrosauridés devaient compter sur leur nombre ou leur vigilance collective plutôt que sur une défense active. Certains hadrosaures possédaient des structures crâniennes (crêtes creuses, boursouflures nasales) qui amplifiaient les vocalisations et permettaient une communication sonore à longue distance au sein du troupeau — aucun crâne de Barsboldia n'étant connu, on ignore s'il disposait d'un tel appareil. Les traces de pas attribuées aux hadrosauridés dans des formations contemporaines montrent souvent des pistes multiples parallèles, indice d'un déplacement en groupe organisé plutôt que solitaire.
La position phylogénétique de Barsboldia a été sujette à révisions successives depuis sa description initiale. Maryańska et Osmólska (1981) l'avaient placé parmi les lambeosaurinés en raison des épines neurales allongées, un caractère jugé à l'époque distinctif de cette sous-famille. Des analyses phylogénétiques postérieures, notamment celles de Prieto-Márquez (2010 et suivantes), ont réévalué cette attribution et rapproché Barsboldia des saurolophines plutôt que des lambeosaurinés. Le problème fondamental reste le même : avec seulement des vertèbres, un bassin partiel et des côtes, les caractères diagnostiques permettant une attribution sous-familiale solide sont peu nombreux. Barsboldia reste donc un hadrosauridé valide mais à position intrafamiliale encore débattue dans la littérature spécialisée.
Parmi les hadrosauridés de grande taille du Crétacé supérieur, Barsboldia se distingue par son contexte géographique asiatique et son matériel fossile particulièrement fragmentaire. Par la taille, il se compare à Saurolophus angustirostris (son contemporain dans le Nemegt) et aux grandes espèces nord-américaines comme Edmontosaurus regalis ou Shantungosaurus giganteus — ce dernier étant probablement le plus grand hadrosauridé connu, avec ses 15 mètres estimés. Les épines neurales allongées de Barsboldia rapprochent sa silhouette de celle d'Ouranosaurus nigeriensis, un hadrosauriforme africain du Crétacé inférieur qui présentait une structure dorsale similaire, bien que les deux animaux ne soient pas étroitement apparentés. Contrairement à Lambeosaurus ou Parasaurolophus, Barsboldia ne semble pas avoir porté de crête céphalique creuse — du moins, aucun matériel crânien attribuable avec certitude à ce genre n'a été trouvé.
Les fossiles de Barsboldia proviennent de la Formation Nemegt, dans le bassin de Nemegt au sud de la Mongolie (province d'Ömnögovi). Les spécimens ont été collectés lors des expéditions polono-mongoles des années 1960-1970, fouilles conjointes qui ont enrichi les collections du Museum of the Earth à Varsovie et de l'Institut de Paléontologie de Mongolie à Oulan-Bator. Le matériel holotype reste limité à des éléments post-crâniens : vertèbres dorsales et sacrées, portions du bassin, côtes. Aucun crâne complet ni membre entier n'a été formellement attribué à ce genre, ce qui contraste fortement avec des contemporains comme Saurolophus angustirostris ou Tarbosaurus bataar, connus par des squelettes relativement complets issus du même gisement. Cette rareté du matériel est en partie inhérente aux conditions de fouille de l'époque et à l'étendue géographique limitée des zones prospectées — les expéditions polono-mongoles couvrant un territoire immense en un temps limité, avec des moyens logistiques contraints. Des missions de prospection plus récentes dans le Nemegt, notamment celles de l'Institut de Paléontologie de Mongolie et de partenaires japonais et américains depuis les années 2000, n'ont pas encore produit de nouveau matériel clairement assignable à Barsboldia. La richesse de la Formation Nemegt reste pourtant considérable, et de nouvelles découvertes pourraient un jour transformer notre compréhension de ce genre énigmatique.
Barsboldia occupe une place modeste dans la culture populaire paléontologique, largement éclipsé par ses voisins du Nemegt plus spectaculaires — Tarbosaurus, Deinocheirus ou Therizinosaurus. Son nom apparaît dans les bases de données paléontologiques de référence et dans les ouvrages encyclopédiques sur les hadrosauridés, mais il ne figure pratiquement jamais dans les documentaires grand public ni dans les collections de jouets ou figurines. Pour les amateurs de dinosaures plus avancés, il incarne surtout les difficultés de classification qui guettent tout genre fondé sur un matériel squelettique incomplet — une leçon récurrente dans l'histoire de la paléontologie des hadrosauridés. Son nom rend hommage à Rinchen Barsbold, paléontologue mongol dont les contributions à la connaissance des dinosaures du Gobi sont immenses et qui mérite d'être bien mieux connu du grand public occidental.
| Période | Crétacé supérieur / Late Cretaceous |
| Ère | Mésozoïque / Mesozoic |
| Âge | 72–70 Ma |
| Localisation | Asie / Asia (Mongolie / Mongolia — Formation Nemegt) |
| Longueur | 10–12 m |
| Hauteur | ~3.5 m |
| Poids | 4 000–7 000 kg |
| Régime | Herbivore |
| Découverte | 1981 |
Barsboldia sicinskii n'est connu que par une colonne vertébrale partielle, un bassin incomplet et des côtes, sans crâne ni membre entier. Décrit en 1981 par Maryańska et Osmólska comme un lambeosauriné à cause de ses épines neurales allongées, il a été rapproché des saurolophines par les analyses de Prieto-Márquez (2010). Sans matériel crânien diagnostique, sa position intrafamiliale reste débattue.
Barsboldia sicinskii était l'un des grands herbivores de la Formation Nemegt en Mongolie, il y a 72 à 70 millions d'années, avec une longueur estimée entre 10 et 12 mètres et un poids de 4 000 à 7 000 kg. Il partageait cet écosystème fluvial avec Saurolophus angustirostris, Deinocheirus mirificus, Therizinosaurus cheloniformis et son prédateur dominant, Tarbosaurus bataar.
Les épines neurales exceptionnellement allongées des vertèbres dorsales de Barsboldia sicinskii — plusieurs fois la hauteur du corps vertébral — sont la principale particularité anatomique connue de ce genre. Leur fonction reste incertaine : voile thermorégulée, bosse graisseuse ou crête musculaire dorsale. Ces structures rappellent superficiellement Ouranosaurus nigeriensis, un hadrosauriforme africain du Crétacé inférieur, sans lien phylogénétique direct.
Les informations de cette fiche sont basées sur des publications scientifiques à comité de lecture.
Reconstitution de Barsboldia sicinskii par Dmitry Bogdanov (2008)
Dmitry Bogdanov, CC BY 3.0, via Wikimedia Commons

Reconstitution de Barsboldia sicinskii par Connor Ashbridge (2023)
Connor Ashbridge (Ddinodan), CC BY-SA 4.0, via Wikimedia Commons